Human Microflora
The human microbiome is a complex ecosystem of microorganisms that plays an important role in digestive health, immune regulation, metabolism, and overall wellness. This educational resource explores vaginal and intestinal microflora, newborn microbial development, and the effects of probiotics on gastrointestinal ecology.
The Vaginal Microflora
Although less complex than the gastrointestinal microflora, the normal vaginal microflora of a premenopausal woman is composed of a variety of bacterial species. Anaerobes are most frequently isolated and appear in numbers of 107 - 109 CFU/ml of vaginal secretion.
Lactobacillus spp. is the most frequently isolated genus found in the highest numbers. They play a role in maintaining the balance of the normal vaginal flora by producing hydrogen peroxide. It has been shown that approximately 70% of premenopausal, healthy women harbor hydrogen peroxide-producing lactobacilli.
Corynebacterium, Staphylococcus and Bacteroides spp. are among the anaerobes frequently isolated.
Intestinal Microflora in the Newborn Infant
Fetuses are sterile in the womb, but beginning with the birth process, infants are exposed to microbes that originate from the mother and the surrounding environment including breast milk or formula.
The infant tends to acquire the flora swallowed from the vaginal fluid at the time of delivery. Because vaginal flora and intestinal flora are similar, an infant's flora may closely mimic the intestinal flora of the mother.
Another factor affecting the intestinal flora of the newborn is delivery mode. A normal vaginal delivery commonly permits transfer of bacteria from the mother to the infant. During cesarean deliveries, this transfer is completely absent.
These infants commonly acquire and are colonized with flora from the hospital environment and therefore their flora may differ from maternal flora. Infants delivered by cesarean section are colonized with more anaerobic bacteria, especially Bacteroides, than vaginally delivered infants.

Delivery mode shapes the initial bacterial inoculum of the newborn. Postnatal factors such as antibiotic use, diet, genetics, and environmental exposure further configure the microbiome during early life.
Reference: “The microbiome in early life: implications for health outcomes.” Tamburini S, et al. Nat Med. 2016 Jul 7;22(7):713-22.
The initial colonizing bacteria vary with the food source of the infant. In breast-fed infants, Bifidobacteria account for more than 90% of the total intestinal bacteria.
The low concentration of protein in human milk, the presence of specific anti-infective proteins such as immunoglobulin A, lactoferrin, lysozyme, and oligosaccharides, as well as production of lactic acid, create an acid milieu and contribute to bifidogenic characteristics.
In bottle-fed infants, Bifidobacteria are not predominant. Instead enterobacteria and gram-negative organisms dominate because of a more alkaline environment and the absence of many prebiotic modulatory factors naturally present in breast milk.
The establishment of intestinal microbial ecology is highly variable at the beginning but gradually becomes more stable and similar to adult microflora by the end of the breastfeeding period.
Other factors affecting intestinal microflora include geographic differences, industrialization, environmental exposure, and administration of antibiotics in neonatal intensive care.
Effects of Probiotics on the Intestinal Microflora
Specific changes may be observed within days after beginning probiotic consumption, depending on dosage and strain capacity to influence gastrointestinal function.
With regular consumption, probiotic bacteria may temporarily colonize the lower intestine. Once intake stops, the number of probiotic microorganisms gradually declines.
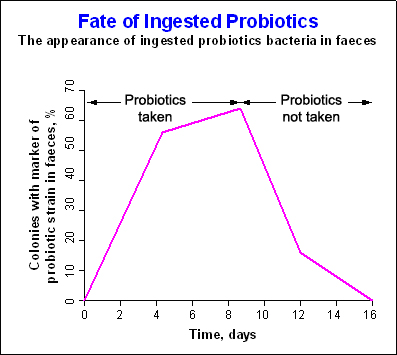
Many studies have demonstrated significant shifts in bacterial counts following consumption of specific probiotic strains, often resulting in increased numbers of Lactobacillus and Bifidobacterium and decreased populations of potentially harmful organisms.
These studies reflect bacterial conditions in fecal matter only and do not fully represent the microbial situation in other regions of the gastrointestinal tract or within the gut mucosal layer.
Probiotic bacteria may also modulate metabolic activity of the gut flora. By lowering intestinal pH, probiotics may interfere with enzymatic activity within the gastrointestinal tract.